视景医疗
知觉学习及其神经机制研究的新进展
1、引言
学习是有机体神经系统的基本功能,它可以改变有机体自身的行为,使得其适应不断变化的外界环境。通常情况下大多数有机体在他们丧失学习机能后就难以适应外界环境的变化。心理学家之所以视知觉信息是因为在人所获得的整个信息中,视觉信息占了绝大多数。因此视知觉学习的研究具有非常重要的理论意义。视知觉学习的主要表现在视觉神经系统从外界环境中提取信息能力的提高。心理学家对于人类视知觉学习的研究已经有较长的历史,并且该领域的研究在最近 30 年逐渐一个前沿和热点问题,许多重要的发现相继被报告出来。知觉学习神经机制的研究主要源自对现象的观察。关于该领域的较早研究主要是从心理物理学的方法入手,研究人员发现了该现象的一些基本特性,如特异性和迁移性等。这些特性的发现启发更多人了解知觉学习的本质,这些发现为探讨其神经生理机制提供了实证依据。
2、知觉学习的实质和特性
人们的知觉随着训练或经验的累积而逐渐改变的现象便是知觉学习的表现形式,这种知觉的改变一旦获得就可以保持相当长的一段时间。然而,训练不达到一定强度才会这种改变一般是不会发生的。Doshe 和 Lu,(1999)提到,一个被广泛认同的结论是特异于训练的特性并可保持较长时间。与其他的学习形式,如认知学习、策略选择等不同,知觉学习都是只改变与训练相关的神经回路。不同于片断记忆,这些记忆大多存储在特定的脑区,但这些脑区并不是和感知觉信号密切相关的(Fahle,2005)。
关于知觉学习的特性,可以概括为以下三点:1)特异性(如:方位、运动方向、视网膜位置、训练任务等):空间频率、训练眼等。特异性又分为刺激特征特异性和任务特异性。2)关于效应的发生时间:一般需要较长时间训练。3)效应持续的时间:从天到年都有报道。被研究人员所普遍关注的一个重要问题便是知觉学习的特异性,这个问题事实上与知觉学习的脑机制是有着密切的关联的。正如 Ahissar 和 Hochstein (1997) 提出的:“知觉学习的特异性和任务的难易程度的有关联的:任务越难,特异性越高;反之亦然”这一假说,他们进一步建立了在本领域影响广泛的逆分级学习模型。详见图 1。
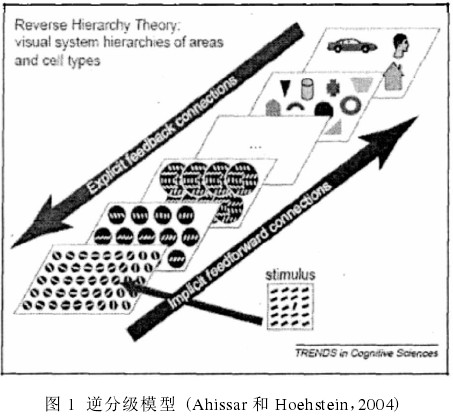 该模型的典型观点是:当信号流不断传播时,从视网膜不同位置及不同方位传递的信息将会逐渐汇聚。所以就会有皮层表征差异越来越弱的情况发生在不同刺激之间。
该模型的典型观点是:当信号流不断传播时,从视网膜不同位置及不同方位传递的信息将会逐渐汇聚。所以就会有皮层表征差异越来越弱的情况发生在不同刺激之间。
3、知觉学习的神经机制
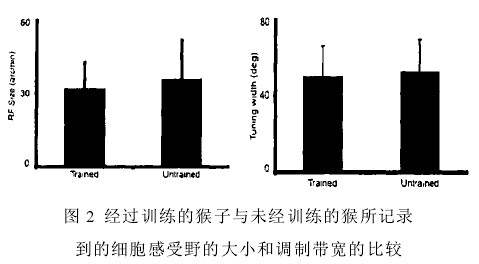
就目前的研究手段来看,电生理技术与行为技术联合应用已经得到了增强,这种趋势大概于 2001 左右起步。我们在这里主要集中于近些年的主要研究成果,大多是一些在猴子上完成的行为学实验的结果。Crist 等人的研究训练猴学习二分任务(bisection),然后用电生理学方法记录了感受野外背景影响特性的变化以及猴子 V1 皮层经典感受野特性以及(Crist et al.,2001)。结果发现: 脑皮层放大系数没有显著性的改变;细胞感受野的电生理学特性,包括其大小及方位选择性(图 2)相比于没有经过训练的猴没有明显的改变。
 Schwartz(2002)等人利用脑成像技术(fMRI)研究了纹理辨别任务知觉学习的神经机制(Sehwartz,Maqueth和Frit-h,2002)。他们发现 V1 皮层同一视网膜投射区域的活动训练眼在显著强于应投射区域非训练眼相的活动。Furmanski等(2004)发现,在 Vl 诱导的 fMRI 信号训练前倾斜方位的光栅显著弱于水平方位光栅所诱导的信号; 但是,这种差异可以经过训练以后被消除。训练的效应最多可以提高 39%,而且行为学测量得到的 detcetion 闪值的提高与这种幅度显著相关。
Schwartz(2002)等人利用脑成像技术(fMRI)研究了纹理辨别任务知觉学习的神经机制(Sehwartz,Maqueth和Frit-h,2002)。他们发现 V1 皮层同一视网膜投射区域的活动训练眼在显著强于应投射区域非训练眼相的活动。Furmanski等(2004)发现,在 Vl 诱导的 fMRI 信号训练前倾斜方位的光栅显著弱于水平方位光栅所诱导的信号; 但是,这种差异可以经过训练以后被消除。训练的效应最多可以提高 39%,而且行为学测量得到的 detcetion 闪值的提高与这种幅度显著相关。
4、总结
有关知觉学习实质、神经生理机制等基本问题上,近些年来许多研究者取得了显著的进展,这两个重要的基本问题可以说得到了一些初步的答案。关于影响知觉学习的因素,人们已经认识到不仅是刺激,而且任务也是一个显著相关的因素。然而,人们对于知觉学习机制的研究尚未形成一致结论:脑成像研究倾向于知觉学习发生在较低阶段;电生理研究则相对支持较高级学习理论。因此一定要综合各种研究手段以做出正确的判断,包括心理物理学与生理学、行为学、脑成像技术,甚至要结合组织化学、分子生物学、药理学、形态学、等手段。将心理物理,行为学和电生理技术结合起来的研究现在己经不少见了,并也取得了不少成果。今后知觉学习研究的主流将是这种多学科之间的联合研究。
作者:曲阜师范大学心理学系王凯
学习是有机体神经系统的基本功能,它可以改变有机体自身的行为,使得其适应不断变化的外界环境。通常情况下大多数有机体在他们丧失学习机能后就难以适应外界环境的变化。心理学家之所以视知觉信息是因为在人所获得的整个信息中,视觉信息占了绝大多数。因此视知觉学习的研究具有非常重要的理论意义。视知觉学习的主要表现在视觉神经系统从外界环境中提取信息能力的提高。心理学家对于人类视知觉学习的研究已经有较长的历史,并且该领域的研究在最近 30 年逐渐一个前沿和热点问题,许多重要的发现相继被报告出来。知觉学习神经机制的研究主要源自对现象的观察。关于该领域的较早研究主要是从心理物理学的方法入手,研究人员发现了该现象的一些基本特性,如特异性和迁移性等。这些特性的发现启发更多人了解知觉学习的本质,这些发现为探讨其神经生理机制提供了实证依据。
2、知觉学习的实质和特性
人们的知觉随着训练或经验的累积而逐渐改变的现象便是知觉学习的表现形式,这种知觉的改变一旦获得就可以保持相当长的一段时间。然而,训练不达到一定强度才会这种改变一般是不会发生的。Doshe 和 Lu,(1999)提到,一个被广泛认同的结论是特异于训练的特性并可保持较长时间。与其他的学习形式,如认知学习、策略选择等不同,知觉学习都是只改变与训练相关的神经回路。不同于片断记忆,这些记忆大多存储在特定的脑区,但这些脑区并不是和感知觉信号密切相关的(Fahle,2005)。
关于知觉学习的特性,可以概括为以下三点:1)特异性(如:方位、运动方向、视网膜位置、训练任务等):空间频率、训练眼等。特异性又分为刺激特征特异性和任务特异性。2)关于效应的发生时间:一般需要较长时间训练。3)效应持续的时间:从天到年都有报道。被研究人员所普遍关注的一个重要问题便是知觉学习的特异性,这个问题事实上与知觉学习的脑机制是有着密切的关联的。正如 Ahissar 和 Hochstein (1997) 提出的:“知觉学习的特异性和任务的难易程度的有关联的:任务越难,特异性越高;反之亦然”这一假说,他们进一步建立了在本领域影响广泛的逆分级学习模型。详见图 1。
3、知觉学习的神经机制
就目前的研究手段来看,电生理技术与行为技术联合应用已经得到了增强,这种趋势大概于 2001 左右起步。我们在这里主要集中于近些年的主要研究成果,大多是一些在猴子上完成的行为学实验的结果。Crist 等人的研究训练猴学习二分任务(bisection),然后用电生理学方法记录了感受野外背景影响特性的变化以及猴子 V1 皮层经典感受野特性以及(Crist et al.,2001)。结果发现: 脑皮层放大系数没有显著性的改变;细胞感受野的电生理学特性,包括其大小及方位选择性(图 2)相比于没有经过训练的猴没有明显的改变。
4、总结
有关知觉学习实质、神经生理机制等基本问题上,近些年来许多研究者取得了显著的进展,这两个重要的基本问题可以说得到了一些初步的答案。关于影响知觉学习的因素,人们已经认识到不仅是刺激,而且任务也是一个显著相关的因素。然而,人们对于知觉学习机制的研究尚未形成一致结论:脑成像研究倾向于知觉学习发生在较低阶段;电生理研究则相对支持较高级学习理论。因此一定要综合各种研究手段以做出正确的判断,包括心理物理学与生理学、行为学、脑成像技术,甚至要结合组织化学、分子生物学、药理学、形态学、等手段。将心理物理,行为学和电生理技术结合起来的研究现在己经不少见了,并也取得了不少成果。今后知觉学习研究的主流将是这种多学科之间的联合研究。
作者:曲阜师范大学心理学系王凯