视景医疗
大脑是怎样处理复杂信息的
引例:
stroop效应(斯特鲁普效应)是指字义对字体颜色的干扰效应。一般认为,念字和说出字体颜色是两个不同的认知过程。Stroop于1935年做了一个实验,他利用的刺激材料在颜色和意义上相矛盾,例如用蓝颜色写“红”这个字,要求被试说出字的颜色,而不是念字的读音,即回答“蓝”。结果发现,说字的颜色时会受到字义的干扰。
麦克劳德(Mecleod)在1991 年总结stroop 效应(斯特鲁普效应)发生机制的5种理论或模型。像早期的相对加工速度理论(赛马理论)、自动化理论、知觉编码理论、Logan 的平行加工模型、平行分布式加工模型。相对加工速度理论出现时间的最早,它认为人们对刺激的两个维度(字词和颜色)加工是平行的,而加工速度不同。读词总快于颜色命名,所以字词首先得到加工。当字词的颜色和颜色信息一致的时候,就会促进对字词的颜色命名,反之对字词的颜色命名则产生干扰。自动化理论受到高度认同的原因,在于它强调区分自动加工和控制加工两个概念,自动加工不需要注意的参与,控制加工则需要有意的控制。在stroop 任务中读词是自动加工,颜色命名是控制加工,所以读词能对颜色命名产生促进或干扰,反之则不会。知觉编码理论强调stroop 的干扰仅发生在知觉编码阶段,加工阶段则不发生。有证据说明stroop的干扰不仅发生在知觉编码阶段,还发生在加工阶段。Logan 的平行加工模型改变过去那种强调加工的系列性,而把stroop 效应看作是收集
证据进行决策的过程。刺激的每个维度的加工速度是由其权重决定的,权重影响每一维度对决策的贡献大小。如果来自某一维度的证据和要求的维度一致,就会降低阈限,从而加快要求维度的加工时间,反之则减慢要求维度的加工速度。平行分布式加工模型(PDP)又称为联结主义和神经网络模型,是对以上几种理论的升华,能解释Stroop 效应中的许多结果。它认为PDP 系统包含很多相互联结的模块,每个模块由许多简单的相互联结的加工单元,每个加工单元负责接收来自其他单元的输入并提供输出。几组模块可以组成通路,每条通路包括一组相互联结的模块。当6D6 系统进行任务操作的时候,就会选择一条通路,通路中的联结组确定了这条通路的强度,从而通路的选择也确定了信息加工的速度和准确性。
而2003 年Robert等提出较新的stroop效应的建构理论。stroop效应的建构理论认为在选择性注意中,基于记忆的两个结构—维度的不平衡性和维度的不确定性引导注意选择那些在刺激维度内或者附近的那些显眼的、奇怪的、或者与任务相关的信息进行加工。
科学家把研究感觉信息处理过程作为揭示脑的奥秘的突破口,其中以视觉系统的研究最为突出。在视知觉的研究中已取得了一系列成果:
1、视网膜的光感受器水平:已克隆出视色素蛋白基因;光电换能过程的第二信使是cGMP(Ca2+),黑暗中,cGMP的作用使Na+通道蛋白活化,导致Na+通道开放,Na+持续内流(暗电流),光感受器细胞去极化;光照引起视色素分解,使视盘膜上的GTP结合蛋白分子活化,后者再激活PDE,迅速分解cGMP,引起Na+通道关闭,暗电流骤降,光感受器细胞膜超极化,这样光能就转化为神经电信号。
2、视网膜,复杂的信息处理(外周脑),研究相当清楚。视网膜这个两维的、多层次信息处理的最后结果,是经由视网膜神经节细胞以动作电位脉冲调频的方式,传递给脑的。
3、感受野(receptor field):视通路中任一神经元都在视网膜(或视野)上有一个代表区域。同心圆拮抗型感受野,包括给光—中心和撤光—中心两类,为心理学马赫带现象提供生理学基础,非同心圆的感受野的细胞对快速运动、运动方向以及某些图形特征产生反应。
4、初级视皮层(纹状皮层),在整个大脑皮层研究最透彻的一部分,面积最大的区域。功能柱:具有相同感受野位置和生理功能的细胞按垂直于皮层表面的柱状结构有序地排列起来。功能柱内细胞具有相同的最优方位、相同的眼优势、相同的最优空间频率。
基本的视觉信息:
1、亮度:可见光的波长范围为380~780nm,亮度是一种外界辐射的物理量在我们视觉中反映出来的心理物理量。亮度定义:光度学计量的一种人的心理物理量,它是在特定的标准条件下,定义一个量值表示辐射量和人平均的光感觉之间的对应关系。人的视知觉对强度和能量的量度,是平均人眼对光辐射量的主观量度。
2、形状:由物体在视觉空间上的亮度分布、颜色分布或运动状态不同而显示出来。对比度(contrast):定义一条亮暗边界的对比度为最大光亮度Lmax与最低亮度Lmin之差除以两者之和,Lmax-Lmin/(Lmax+Lmin)。实验常用的刺激图有光点、光环、光条、光栅、随机点图形,无不涉及图形与背景的对比度测定,因此对比度是一个关键的视觉刺激量。一条视觉上的亮暗边界还有一个重要的属性,即其在二维空间上的朝向——方位,方位选择性已成为公认的形状视觉中最为重要的视觉刺激参数。
3、运动(运动方向和速度):视觉系统对运动目标十分敏感,兔的视网膜中有大量的对运动目标极为敏感的给光-撤光类的W型细胞。
4、颜色:主观感觉,中枢机制尚不清。例如,用适当的红绿兰三种颜色同时刺激,颜色感觉完全消失,只有亮度感觉。在暗视时,只能区别亮度,单变量原理
明视时,三种视锥细胞共同工作,不同波长的光刺激不同的感受系统的相对程度不同,产生颜色的感觉器水平,色觉的三变量性。
5、立体(或深度)视觉:同一物体在左右眼的视网膜象是有微小差别的,这种差别为视觉系统提供了立体视觉(stereopsis)的最基本的信息——视差(disparity )。立体视觉使我们的得到了一个对周围世界的生动而精确的相对的深度分辨。只有在双眼同时运作才有的感觉。
图1是眼球的结构,一束平行光线从眼球的最前方的角膜穿过,经过前房、晶状体、玻璃体,最后聚焦于视网膜上的一点。整个眼球的折光装置类似于一个凸透镜,其中晶状体是可以调节的,根据视物的远近进行调节,光线的强弱可以调节瞳孔的大小。如果视点落在视网膜前,则为近视,反之为远视。一般是通过眼镜来调节折光率,目前有一些治疗近视的技术是处理角膜,改变角膜的折光性。
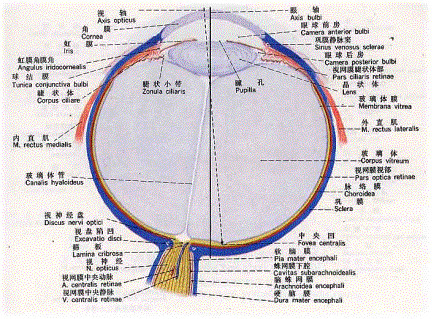
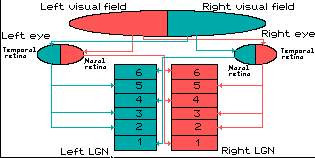
图1 眼球结构 人视网膜可分为以视轴为中心直径约5~6mm的中央区和周边区。中央区又可分为中央凹(fovea)、旁中央凹和远中央凹;周边区也分为近周边区、中周边区和远周边区。 光线落在视网膜上后,被感光细胞接受,产生超极化响应,将光信号转化为电信号。然后通过图2的视觉传导通路传递到脑皮层的视区,形成视觉。视觉传导途径:感受器(视杆和视锥细胞)→第一级神经元(双极细胞)→第二级神经元(节细胞)→视神经→视交叉(鼻侧半纤维交叉,颞侧不交叉)→视束→第三级神经元(外侧膝状体)→视辐射→内囊枕部→枕叶视区。视束中的纤维有部分经上丘臂进入上丘和顶盖前区→组成顶盖脊髓束→参与视、听觉反射。
视觉传导通路的不同部位损伤所引起的视野的变化:
a.一侧视神经损伤→患侧视野全盲
b.视交叉中央部(交叉纤维)损伤→双侧视野颞侧偏盲
c.视交叉外侧部损伤→患侧视野鼻侧偏盲
d.一侧视束视辐射或视觉中枢损伤→双眼视野对侧同向性偏盲(即患侧视野鼻侧半偏盲和健侧视野颞侧半偏盲)
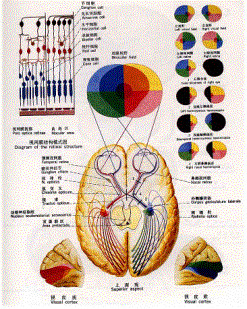
图2 视觉通路 视网膜,又称为外周脑,从起源来说与大脑相同,是与外界有直接联系的部分。从组织上来讲,包括十层细胞,它们构成了一个复杂的细胞网络,具有初步的信息处理功能。图3是视网膜的简化图。感受器细胞(感光细胞,receptor cell, RC)将光量子能量转换成电信号,具体地说就是光刺激变成感受器细胞的膜电位超极化,(光致超极化效应),经化学突触将信号传到双极细胞,双极细胞进而又将信号处理后经化学突触传递到神经节细胞,神经节细胞是唯一的能将视网膜处理后的视觉信息编码为神经冲动传输到脑的细胞。介于感光细胞和双极细胞之间有一水平细胞层,从光感受器接收信息,并反馈输出到光感受器,同时也输出到双极细胞,在这三种细胞间形成了复杂的突触联系网络层,作为外网状层。内网状层,双极细胞——无足细胞层——神经节细胞层。网间细胞接受无足细胞的输入,逆行投射到外网状层的水平细胞形成突触,偶尔也与双极细胞形成突触,在内网状层与外网状层之间形成了一条离心反馈通路。
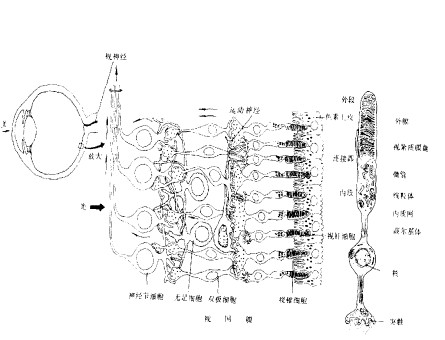
图3 视网膜结构 感受器细胞包括外段(outer segment,OS)(形状有的呈杆状,有的呈锥状)和内段(inner segment, IS),中间为一个细的连接颈。外段充满了由膜围成扁囊状结构,在膜上镶嵌有数以百万计的视色素(visiual pigment, VP),由视蛋白和视黄醛构成,两者的差异在于视蛋白的不同。感受器细胞分类两类:视锥细胞和视杆细胞。
视锥细胞(core,C): 6.5百万/单眼,光敏感度低,强光刺激才能引起兴奋,但具有分辨颜色的能力。中央凹,仅视锥细胞,密度最高,约150000个/mm2。中央凹的结构特点均为特高的视锐度创造了条件,它是灵长类视网膜适应高视锐度的需要而分化的结果。视觉最敏感。鸽子只有视锥细胞。3种视锥细胞,包含不同的视紫蓝质分子,绿视锥细胞 450~675nm,红-蓝。530nm,绿光。蓝视锥细胞,455nm(蓝光);红视锥细胞,625nm(橙色光)。
视杆细胞(rod, R):1.25亿/单眼,视紫红质,对弱光敏感,一个光量子可引起一个细胞兴奋,5个光量子就可使人眼感觉到一个闪光,不能分辨颜色。猫头鹰只有视杆细胞。
光照,视紫红质中的顺式视黄醛变构成全反式视黄醛,视蛋白与之分离,视黄醛在酶作用下还原成Va,在暗处,在酶作用下由全反式生成顺式。构象变化激活了转导蛋白(T)一个光量子所激活的视紫红质分子能与约500个转蛋白的分子相互作用,使信号放大,转导蛋白转而激活磷酸二酯酶(PDE),PDE又使cGMP降解为非活性的GMP,一个PDE分子每秒钟可使2000个cGMP分子分解,cGMP含量的下降,造成了Na+不能再流入细胞内,于是此细胞电位变得更负,超极化的视杆细胞不再继续释放神经递质,递质释放量下降,无论刺激多强,只能给出分级的超极化电位,不产生动作电位(无冲动神经元),经过这一系列级联反应,一个光量子信号放大了约1亿倍。
双极细胞(bipolar cell, BC):只能给出分级电位,不产生动作电位。明显的呈现中心和周边同心圆拮抗方式。对感受野中心的光刺激呈去极化,给光——中心双极细胞;对中心光照呈超极化反应,超极化或撤光—中心双极细胞。
色拮抗双极细胞,单拮抗细胞:感受野中心对红光最敏感,周边区对绿光最敏感。(心理学)时间色对比现象的神经基础,在注视红色一段时间后,突然观看一张白纸,会感到绿色出现的现象,反之亦然。双拮抗细胞,中心区和周边区刺激波长改变时,反应的极性也会翻转,同时色对比现象,当一灰色区域被一红色区域包围时,灰色区域呈现出绿色,反之亦然。
神经节细胞(ganglion cell, GC):同心圆拮抗式感受野(视系统中的单细胞活动,若受一定的时间和空间构型的光刺激,视网膜某区域而调制时,该区域就称为该细胞的感受野)同心圆拮抗形式,即感受野一般是由中心的兴奋区和周边抑制区所组成的同心圆结构,在功能上是相互拮抗的(图4)。给光区域:给光时,GC单细胞发放频率升高;撤光区域:撤光时,GC单细胞发放频率升高;on-off:给光、撤光均升高。1965年,Rodieck关于同心圆拮抗式感受野的数学模型高斯分布的性质,高斯差模型。(difference of two Gaussians) 。神经节细胞的同心圆拮抗式感受野可以解释心理学中著名的马赫带(Mach band)现象,马赫是19世纪奥地利著名的物理学家:在观察一个亮度渐变的边缘时,发现主观感觉在亮的一端呈现一个特别亮的亮带,在暗的一端呈现一个特别暗的暗带。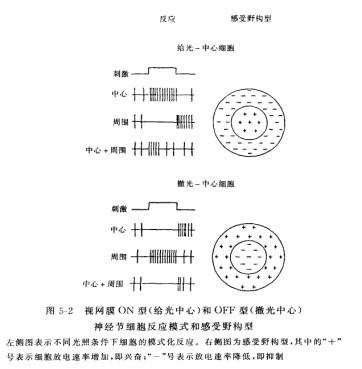
图4 神经节细胞的感受野 感受器细胞的总数是视网膜节细胞的100倍,外膝体神经元则与神经节细胞数目几乎相等,视皮层17区第4层的细胞数几乎为外膝体细胞数的40倍。所以在17区的第4层,即视皮层的信息入口处存在很大的信息处理容量,从而为视皮层内第一级的精细信息加工创造了条件。
外膝状体核(LGN):丘脑感觉核中的一个,专司视觉信 息处理,规则的排列为六层弯曲的结构,上部四层内细胞较小,称小细胞层(parvocellular layers, P-层),下部二层内细胞较大,称大细胞层(magnocellular layers, M-层)。
 1、(猴)对单侧外膝状体核来说,其1,4,6层只接受对侧眼(鼻侧)的视网膜来的投射输入,而2,3,5层仅接受同侧眼(颞侧)的视网膜来的投射输入,单侧外膝状体只能得到双眼输入的对侧视野内的视觉信息。
1、(猴)对单侧外膝状体核来说,其1,4,6层只接受对侧眼(鼻侧)的视网膜来的投射输入,而2,3,5层仅接受同侧眼(颞侧)的视网膜来的投射输入,单侧外膝状体只能得到双眼输入的对侧视野内的视觉信息。
2、来自视网膜相应点的神经节细胞轴突,投射到外膝状体核各层时是极有规律的,如将外膝状体各层接收投射的响应细胞部位连接起来, 就会得到大体上与各层边界垂直的线,称作投射线;
3、视网膜中央区细胞在外膝状体占有的投射区面积要比视网膜边缘区细胞的投射区大得多,因为在视网膜中央区,各类视网膜细胞密度最高。
 视皮层,现知与视觉有关的大脑皮层多达35个,自皮层表面到白质分成6层(图9),外膝状体核处理后的视觉信息首先传到皮层17区,(I区或纹状体区)。外膝状体细胞轴突末梢终止于第4层内,然而再与2,3层细胞,第5,6层细胞建立突触联系。V1:纹状皮层(17区),V2 第2视区, MST, 内侧上颞区, MT,中央颞区。细胞类型有星形细胞(stellate cell)和锥体细胞(pyramidal cell)。
视皮层,现知与视觉有关的大脑皮层多达35个,自皮层表面到白质分成6层(图9),外膝状体核处理后的视觉信息首先传到皮层17区,(I区或纹状体区)。外膝状体细胞轴突末梢终止于第4层内,然而再与2,3层细胞,第5,6层细胞建立突触联系。V1:纹状皮层(17区),V2 第2视区, MST, 内侧上颞区, MT,中央颞区。细胞类型有星形细胞(stellate cell)和锥体细胞(pyramidal cell)。
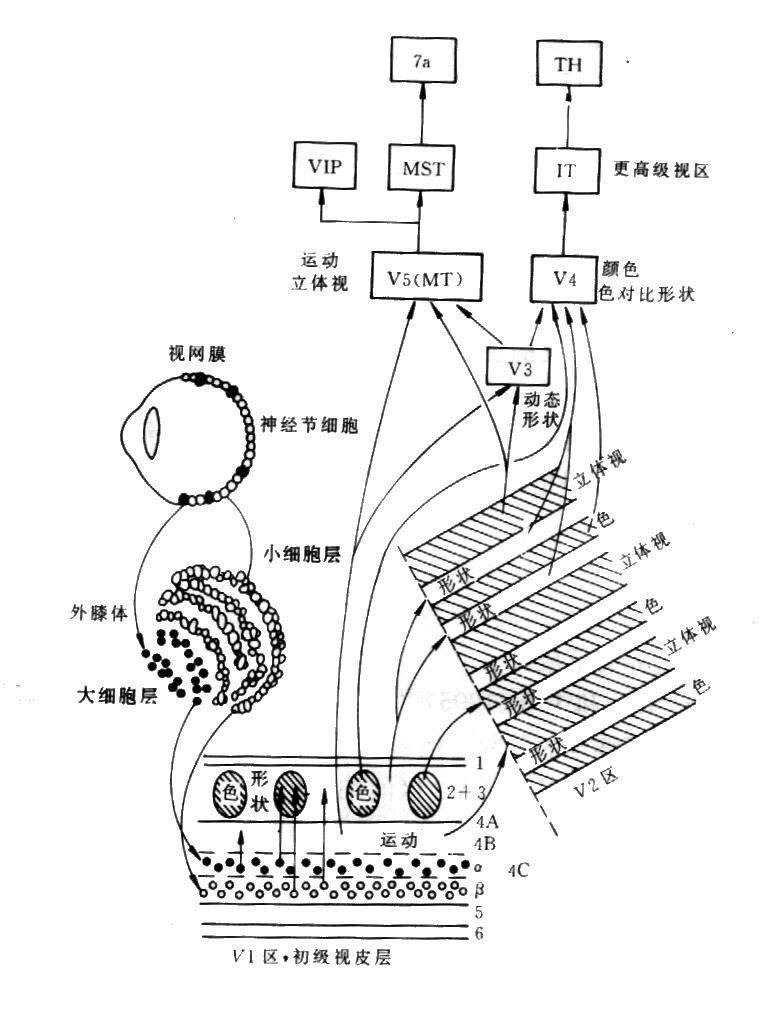
图9 视皮层 大脑皮层17区即视觉初级皮层或视1区,是大脑皮层中被研究得最透彻的区域。当Hubel和Wiesel首次研究视皮层细胞对光刺激的反映时,意外地发现这些细胞都有共同的特点,即对大面积弥散光刺激没有反应,而对有一定方位或朝向的亮暗对比边或光棒、暗棒有强烈反应,若该刺激物的方位偏离该细胞"偏爱"的最优方位,细胞反应便停止或骤减。因此,强烈的方位选择性是绝大多数视皮层细胞的共性。
具体地说视皮层17区和18区的细胞可分为简单细胞(simple cells)和复杂细胞(complex cells)两大类。简单细胞主要分布在视皮层17区的第4层内,感受野较小,呈狭长形,用小光点可以测定,对大面积的弥散光无反应,而对处于拮抗区边缘一定方位和一定宽度的条形刺激有强烈的反应,因此比较适合于检测具有明暗对比的直边,对边缘的位置和方位有严格的选择性,对每一个简单细胞,都有一个最有方位,在此方位上细胞反应最强烈。
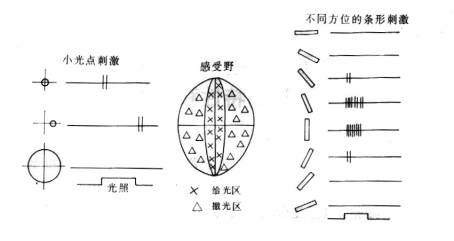
图10 简单细胞的方位选择性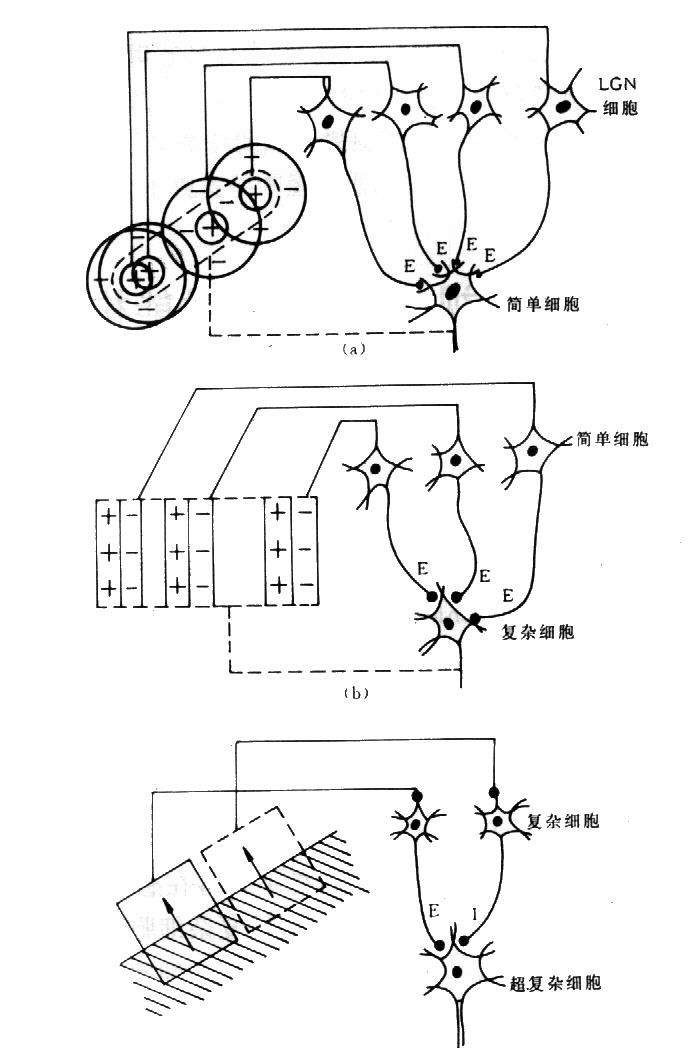
图11 复杂细胞 复杂细胞同样处在要求刺激具有特定的方位,但对其在感受野中的位置无严格要求。多分布在皮层17区(占大部分细胞)和18区,在19区很少看到。形态学上可能是第3和第5层中的锥体细胞。超复杂细胞对条形刺激的反应类似复杂细胞,不同之处是超复杂细胞感受野的一端或两端有很强的抑制区,因此要求条形刺激有一定长度,过长时就产生抑制,反应减少或消失。
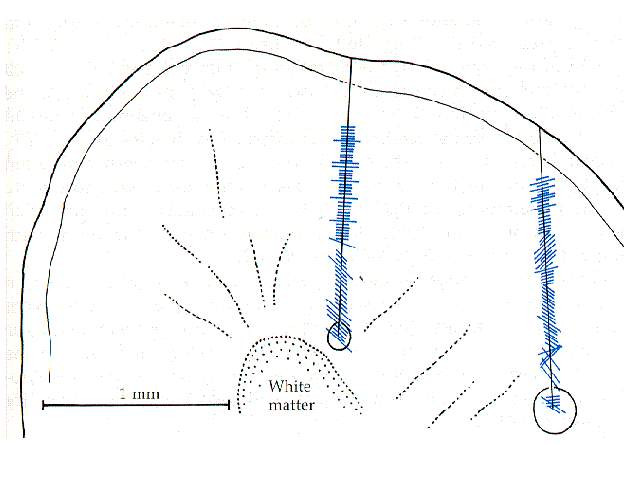
图12 功能柱发现实验 Hubel和Wiesel用单细胞的微电极纪录结合某些特殊的组织学技术,发现许多具有相同视觉功能特性的皮层细胞,在视皮层上按一定的规则(空间上的结构)排列起来,这种按功能排列的皮层结构——称为皮层的功能构筑(functional architecture )。他们从1962年开始研究,1981年获得诺贝尔医学和生理学奖。
垂直和倾斜穿刺连续纪录得到的细胞最优方位分布,短线的长度代表该细胞反应的强弱,短线的朝向代表每一纪录到细胞的最优方位。当用微电极以垂直于视皮层表面的方向插入时,由浅入深地依次纪录到的各类细胞的感受野再视网膜上的位置差不多都是重叠的,而且都有几乎相同的最优方位。如果以微电极倾斜方向穿刺视皮层,所纪录到的各个细胞不仅感受野位置连续的发生漂移,而且他们的最优方位就大致上以10度/50um的变化绿,按顺时针或逆时针的方向发生连续变化,有时在旋转90~270度以后,旋转方向发生逆转。因此,Hubel和Wiesel设想具有相同最优方位的视皮层细胞,是柱状垂直于皮层表面排列的,而且是连成薄片状的薄层所组成,故如要包括180全方位的这些薄板的总宽度应是0.9mm左右,这样一套薄板结构称为方位柱(orientation column)。眼优势柱:大多数双眼细胞接受双眼输入时,总是有一侧眼占优势的,眼优势决定于交叉和未交叉视通道激活4C层细胞的比例,可以根据分别刺激同侧或对侧眼的感受野所产生反应的大小来决定。眼优势柱与方位柱是相对独立的功能结构系统,他们既不平行又不成直角,而是随即交叉的。空间频率柱,远不如方位柱和眼优势柱那样界限分明。
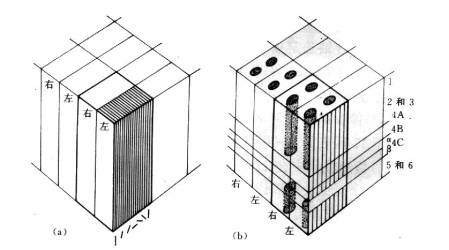
图13 功能柱 纹状皮层(striate cortex),70年代初期提出了视觉皮层功能专门化的概念,认为颜色、形状、运动及可见世界的其他属性都是分别处理的。PET研究,发现视力正常的人看一副蒙德里安水彩风景画(一种毫无可识别物的抽象景色)时,区域性大脑血流增加最大是发生在V4,如果看动着的黑白方块时,最大的血流量是发生在一个与V4完全分隔开的更靠近侧面的区域中V5。这为人类视觉皮层的功能专门化特征提供了直接证据,同时表明在这两种刺激条件下,V1区(或许还有相邻的V2)也呈现过区域性大脑血流显著增加的现象。V1区通常是富含细胞层的,染色后所具有的特征是一些细胞柱从皮层表面延伸到底下叫做白质的神经组织,如果从平行与皮层表面的切面中去看,这些细胞柱就像是重度染色的色斑或蓬松的团块,彼此间被一些轻度染色的色斑间区隔开。发现波长选择型细胞是集中在V1的色斑里,而形状选择型细胞则是集中在色斑间区里。
在V2区,构筑物的形状是一些粗细条纹,彼此间被一些轻度染色的中间条纹分开,对波长选择型细胞是集中在细条纹中,对方向性运动有选择性的细胞则是存在于粗条纹中,对形状敏感的细胞则是粗条纹和中间条纹中都有所分布。这些事实是我们可以勾画出四个与不同视觉特征有关的并行系统:一个是针对运动的,一个是针对颜色的,两个是针对形状的。两个计算上大不相同的系统是运动系统和色彩系统,对于运动系统来说,关键性纹状前区是V5,其输入信号的传递是从视网膜经外侧膝状核的大细胞层到V1的4B层,再以直接和通过V2的粗条纹两种方式从那里传送到V5。色彩系统是依靠V4区,其输入信号经外侧膝状核的细胞层传递给V1的色斑,然后以直接和通过V2的细条纹两种方式前往V4。
特定皮层区的损害会相应的造成特有的视觉综合症:V4区的损害曾导致过全色盲(只能看到灰色梯度),不同与单纯色盲,不但不能看见或认识彩色世界,甚至连罹患这种疾病之前所曾见过的色彩是什么样子都回忆不起来。如果它们的视网膜和V1区域都很正常,它们对形状、深度及运动的了解就仍是完好的。V5区的损害会造成运动盲(Akinetopsia),既看不见又不能理解运动中的世界,当物体处于静止状态时,他们可能完全看得见,但是与之相关的运动却会使物体消失,视觉的其他特征却依然没有受损。假定形状和颜色是在皮层内分离的,但是,从来没有人报道过形状是觉得完全丧失和特定丧失。17区不投射到对侧皮层而投射到18,19区和别的皮层区域,18区向前投射到19区,此外,18,19区又投射到对侧皮层对称点和颞叶中部和下部一些区域。
根据以上对视网膜、外膝体和初级视皮层的研究成果,我们可以看到视觉信息处理机制是既平行又分级串行的信息处理机制,如图9。视系统组织成不同的通路对视觉信息的不同侧面进行传递和处理。从外侧膝状体不同层细胞投射至视皮层至少有3条通路,分别对运动、空间(包括立体视觉)信息、形状、图像信息以及颜色信息进行分析处理这三条通路并非完全独立,在不同水平上存在着交叉性连接。单个细胞或细胞群在感知的水平上并不表示某种特征状态,而仅仅表示被感知物的某些特殊的方面。换言之,分离的个部分所表示的并非整体,而使它们间的关系才构成整体的感知。在这种综合的过程中,必须要有“注意”(attention)的参与,“注意”会强调某一物体的特异的性质,突出对生存和生活有重要意义的视觉目标,同时又忽略该物体的其它特性和其他物体。视觉信息的整合是一个过程,在这个过程中对视觉世界的感知和理解是同时发生的。
更高层次的脑信息处理机制,目前提出一些假设,主要有:
1、“主教细胞”假说:根据Hubel和Wiesel提出的分级假说(hierachical hypothesis),就可以推导是否存在祖母细胞。所存在的关键问题是(1)组合爆炸;(2) 信息的集成问题。
2、细胞群假设 :由一群细胞的时空发放模式来反映刺激的类型。问题是(1) 重叠灾难 (2) 难以表达“等级结构”(3)特征捆绑问题。
3、时空编码理论 :动态细胞群。1989年,C.M. Gray & W. Singer等发现了视皮层神经元活动的gamma(40Hz)同步振荡的研究结果,引起了神经科学界的极大关注。1995年,提出了时空编码理论,主要有以下内容:(1) 是建立在神经元脉冲发放的精细时间结构上的,与同步振荡不同的是,脉冲不是等间隔的振荡脉冲,脉冲的间隔也被用于编码信息(2) 神经元是作为脉冲时间一致性检测器来工作的,也就是说,一个神经元自身的脉冲发放情况,由送入该元的输入脉冲在时间上的一致性来确定的,比如说接近同时到达的脉冲数超过一定阈值,则该元发放。与传统神经网络的整合-放电单元不同,通过一致性检测单元传递的不是发放率,而是脉冲发放时间;(3) 通过局部的一致性检测,形成“动态细胞群”。一致性检测是局部事件,动态细胞群则是系统级事件,是一群细胞通过脉冲发放时间临时链接到一起形成的,注意,并不要求这些细胞的脉冲发放是同步的,只要相关就可以了。(4) 相关的关系是根据上下文(任务)形成的,细胞群可以根据上下文的变化进行快速地重组。之所以称为时-空编码,“空”是指空间连接,是一种硬件的拓朴结构,就是神经元间的物理连接;“时”则是指在硬件结构上传递的时间模式,这种模式可以决定动态的连接结构,这种时间模式的产生与硬件连接有关,但时间变量提供了传递信息的巨大能力,结果是可以在同一硬件上形成许多不同的“动态结构”。“动态细胞群”是编码信息的基本单位,是和任务有关的、以自涌方式形成的细胞的瞬时组合。“动态细胞集群”假设可以有效地解决“重叠灾难”问题、捆绑问题、上下文关系表达问题。
对视觉信息处理的根本问题是:
1、 脑的各部分越来越趋向于分工处理各种各样的视觉特征,然后我们的视觉却把各种视觉特征综合成一个完整的视觉实体。脑象——〉心理象、认知象?脑活动——〉精神活动?
2、 视觉图像如何在脑中贮存和利用。在梦中,转动头部探望侧面或背面的物体,视野中的景象在视网膜上的光学象也朝与头部运动相反的方向移动,但看到的外界景象却纹丝不同,脑似乎有补偿机制。
stroop效应(斯特鲁普效应)是指字义对字体颜色的干扰效应。一般认为,念字和说出字体颜色是两个不同的认知过程。Stroop于1935年做了一个实验,他利用的刺激材料在颜色和意义上相矛盾,例如用蓝颜色写“红”这个字,要求被试说出字的颜色,而不是念字的读音,即回答“蓝”。结果发现,说字的颜色时会受到字义的干扰。
麦克劳德(Mecleod)在1991 年总结stroop 效应(斯特鲁普效应)发生机制的5种理论或模型。像早期的相对加工速度理论(赛马理论)、自动化理论、知觉编码理论、Logan 的平行加工模型、平行分布式加工模型。相对加工速度理论出现时间的最早,它认为人们对刺激的两个维度(字词和颜色)加工是平行的,而加工速度不同。读词总快于颜色命名,所以字词首先得到加工。当字词的颜色和颜色信息一致的时候,就会促进对字词的颜色命名,反之对字词的颜色命名则产生干扰。自动化理论受到高度认同的原因,在于它强调区分自动加工和控制加工两个概念,自动加工不需要注意的参与,控制加工则需要有意的控制。在stroop 任务中读词是自动加工,颜色命名是控制加工,所以读词能对颜色命名产生促进或干扰,反之则不会。知觉编码理论强调stroop 的干扰仅发生在知觉编码阶段,加工阶段则不发生。有证据说明stroop的干扰不仅发生在知觉编码阶段,还发生在加工阶段。Logan 的平行加工模型改变过去那种强调加工的系列性,而把stroop 效应看作是收集
证据进行决策的过程。刺激的每个维度的加工速度是由其权重决定的,权重影响每一维度对决策的贡献大小。如果来自某一维度的证据和要求的维度一致,就会降低阈限,从而加快要求维度的加工时间,反之则减慢要求维度的加工速度。平行分布式加工模型(PDP)又称为联结主义和神经网络模型,是对以上几种理论的升华,能解释Stroop 效应中的许多结果。它认为PDP 系统包含很多相互联结的模块,每个模块由许多简单的相互联结的加工单元,每个加工单元负责接收来自其他单元的输入并提供输出。几组模块可以组成通路,每条通路包括一组相互联结的模块。当6D6 系统进行任务操作的时候,就会选择一条通路,通路中的联结组确定了这条通路的强度,从而通路的选择也确定了信息加工的速度和准确性。
而2003 年Robert等提出较新的stroop效应的建构理论。stroop效应的建构理论认为在选择性注意中,基于记忆的两个结构—维度的不平衡性和维度的不确定性引导注意选择那些在刺激维度内或者附近的那些显眼的、奇怪的、或者与任务相关的信息进行加工。
科学家把研究感觉信息处理过程作为揭示脑的奥秘的突破口,其中以视觉系统的研究最为突出。在视知觉的研究中已取得了一系列成果:
1、视网膜的光感受器水平:已克隆出视色素蛋白基因;光电换能过程的第二信使是cGMP(Ca2+),黑暗中,cGMP的作用使Na+通道蛋白活化,导致Na+通道开放,Na+持续内流(暗电流),光感受器细胞去极化;光照引起视色素分解,使视盘膜上的GTP结合蛋白分子活化,后者再激活PDE,迅速分解cGMP,引起Na+通道关闭,暗电流骤降,光感受器细胞膜超极化,这样光能就转化为神经电信号。
2、视网膜,复杂的信息处理(外周脑),研究相当清楚。视网膜这个两维的、多层次信息处理的最后结果,是经由视网膜神经节细胞以动作电位脉冲调频的方式,传递给脑的。
3、感受野(receptor field):视通路中任一神经元都在视网膜(或视野)上有一个代表区域。同心圆拮抗型感受野,包括给光—中心和撤光—中心两类,为心理学马赫带现象提供生理学基础,非同心圆的感受野的细胞对快速运动、运动方向以及某些图形特征产生反应。
4、初级视皮层(纹状皮层),在整个大脑皮层研究最透彻的一部分,面积最大的区域。功能柱:具有相同感受野位置和生理功能的细胞按垂直于皮层表面的柱状结构有序地排列起来。功能柱内细胞具有相同的最优方位、相同的眼优势、相同的最优空间频率。
基本的视觉信息:
1、亮度:可见光的波长范围为380~780nm,亮度是一种外界辐射的物理量在我们视觉中反映出来的心理物理量。亮度定义:光度学计量的一种人的心理物理量,它是在特定的标准条件下,定义一个量值表示辐射量和人平均的光感觉之间的对应关系。人的视知觉对强度和能量的量度,是平均人眼对光辐射量的主观量度。
2、形状:由物体在视觉空间上的亮度分布、颜色分布或运动状态不同而显示出来。对比度(contrast):定义一条亮暗边界的对比度为最大光亮度Lmax与最低亮度Lmin之差除以两者之和,Lmax-Lmin/(Lmax+Lmin)。实验常用的刺激图有光点、光环、光条、光栅、随机点图形,无不涉及图形与背景的对比度测定,因此对比度是一个关键的视觉刺激量。一条视觉上的亮暗边界还有一个重要的属性,即其在二维空间上的朝向——方位,方位选择性已成为公认的形状视觉中最为重要的视觉刺激参数。
3、运动(运动方向和速度):视觉系统对运动目标十分敏感,兔的视网膜中有大量的对运动目标极为敏感的给光-撤光类的W型细胞。
4、颜色:主观感觉,中枢机制尚不清。例如,用适当的红绿兰三种颜色同时刺激,颜色感觉完全消失,只有亮度感觉。在暗视时,只能区别亮度,单变量原理
明视时,三种视锥细胞共同工作,不同波长的光刺激不同的感受系统的相对程度不同,产生颜色的感觉器水平,色觉的三变量性。
5、立体(或深度)视觉:同一物体在左右眼的视网膜象是有微小差别的,这种差别为视觉系统提供了立体视觉(stereopsis)的最基本的信息——视差(disparity )。立体视觉使我们的得到了一个对周围世界的生动而精确的相对的深度分辨。只有在双眼同时运作才有的感觉。
图1是眼球的结构,一束平行光线从眼球的最前方的角膜穿过,经过前房、晶状体、玻璃体,最后聚焦于视网膜上的一点。整个眼球的折光装置类似于一个凸透镜,其中晶状体是可以调节的,根据视物的远近进行调节,光线的强弱可以调节瞳孔的大小。如果视点落在视网膜前,则为近视,反之为远视。一般是通过眼镜来调节折光率,目前有一些治疗近视的技术是处理角膜,改变角膜的折光性。
图1 眼球结构
视觉传导通路的不同部位损伤所引起的视野的变化:
a.一侧视神经损伤→患侧视野全盲
b.视交叉中央部(交叉纤维)损伤→双侧视野颞侧偏盲
c.视交叉外侧部损伤→患侧视野鼻侧偏盲
d.一侧视束视辐射或视觉中枢损伤→双眼视野对侧同向性偏盲(即患侧视野鼻侧半偏盲和健侧视野颞侧半偏盲)
图2 视觉通路
图3 视网膜结构
视锥细胞(core,C): 6.5百万/单眼,光敏感度低,强光刺激才能引起兴奋,但具有分辨颜色的能力。中央凹,仅视锥细胞,密度最高,约150000个/mm2。中央凹的结构特点均为特高的视锐度创造了条件,它是灵长类视网膜适应高视锐度的需要而分化的结果。视觉最敏感。鸽子只有视锥细胞。3种视锥细胞,包含不同的视紫蓝质分子,绿视锥细胞 450~675nm,红-蓝。530nm,绿光。蓝视锥细胞,455nm(蓝光);红视锥细胞,625nm(橙色光)。
视杆细胞(rod, R):1.25亿/单眼,视紫红质,对弱光敏感,一个光量子可引起一个细胞兴奋,5个光量子就可使人眼感觉到一个闪光,不能分辨颜色。猫头鹰只有视杆细胞。
光照,视紫红质中的顺式视黄醛变构成全反式视黄醛,视蛋白与之分离,视黄醛在酶作用下还原成Va,在暗处,在酶作用下由全反式生成顺式。构象变化激活了转导蛋白(T)一个光量子所激活的视紫红质分子能与约500个转蛋白的分子相互作用,使信号放大,转导蛋白转而激活磷酸二酯酶(PDE),PDE又使cGMP降解为非活性的GMP,一个PDE分子每秒钟可使2000个cGMP分子分解,cGMP含量的下降,造成了Na+不能再流入细胞内,于是此细胞电位变得更负,超极化的视杆细胞不再继续释放神经递质,递质释放量下降,无论刺激多强,只能给出分级的超极化电位,不产生动作电位(无冲动神经元),经过这一系列级联反应,一个光量子信号放大了约1亿倍。
双极细胞(bipolar cell, BC):只能给出分级电位,不产生动作电位。明显的呈现中心和周边同心圆拮抗方式。对感受野中心的光刺激呈去极化,给光——中心双极细胞;对中心光照呈超极化反应,超极化或撤光—中心双极细胞。
色拮抗双极细胞,单拮抗细胞:感受野中心对红光最敏感,周边区对绿光最敏感。(心理学)时间色对比现象的神经基础,在注视红色一段时间后,突然观看一张白纸,会感到绿色出现的现象,反之亦然。双拮抗细胞,中心区和周边区刺激波长改变时,反应的极性也会翻转,同时色对比现象,当一灰色区域被一红色区域包围时,灰色区域呈现出绿色,反之亦然。
神经节细胞(ganglion cell, GC):同心圆拮抗式感受野(视系统中的单细胞活动,若受一定的时间和空间构型的光刺激,视网膜某区域而调制时,该区域就称为该细胞的感受野)同心圆拮抗形式,即感受野一般是由中心的兴奋区和周边抑制区所组成的同心圆结构,在功能上是相互拮抗的(图4)。给光区域:给光时,GC单细胞发放频率升高;撤光区域:撤光时,GC单细胞发放频率升高;on-off:给光、撤光均升高。1965年,Rodieck关于同心圆拮抗式感受野的数学模型高斯分布的性质,高斯差模型。(difference of two Gaussians) 。神经节细胞的同心圆拮抗式感受野可以解释心理学中著名的马赫带(Mach band)现象,马赫是19世纪奥地利著名的物理学家:在观察一个亮度渐变的边缘时,发现主观感觉在亮的一端呈现一个特别亮的亮带,在暗的一端呈现一个特别暗的暗带。
图4 神经节细胞的感受野
外膝状体核(LGN):丘脑感觉核中的一个,专司视觉信 息处理,规则的排列为六层弯曲的结构,上部四层内细胞较小,称小细胞层(parvocellular layers, P-层),下部二层内细胞较大,称大细胞层(magnocellular layers, M-层)。
2、来自视网膜相应点的神经节细胞轴突,投射到外膝状体核各层时是极有规律的,如将外膝状体各层接收投射的响应细胞部位连接起来, 就会得到大体上与各层边界垂直的线,称作投射线;
3、视网膜中央区细胞在外膝状体占有的投射区面积要比视网膜边缘区细胞的投射区大得多,因为在视网膜中央区,各类视网膜细胞密度最高。
图9 视皮层
具体地说视皮层17区和18区的细胞可分为简单细胞(simple cells)和复杂细胞(complex cells)两大类。简单细胞主要分布在视皮层17区的第4层内,感受野较小,呈狭长形,用小光点可以测定,对大面积的弥散光无反应,而对处于拮抗区边缘一定方位和一定宽度的条形刺激有强烈的反应,因此比较适合于检测具有明暗对比的直边,对边缘的位置和方位有严格的选择性,对每一个简单细胞,都有一个最有方位,在此方位上细胞反应最强烈。
图10 简单细胞的方位选择性
图11 复杂细胞
图12 功能柱发现实验
垂直和倾斜穿刺连续纪录得到的细胞最优方位分布,短线的长度代表该细胞反应的强弱,短线的朝向代表每一纪录到细胞的最优方位。当用微电极以垂直于视皮层表面的方向插入时,由浅入深地依次纪录到的各类细胞的感受野再视网膜上的位置差不多都是重叠的,而且都有几乎相同的最优方位。如果以微电极倾斜方向穿刺视皮层,所纪录到的各个细胞不仅感受野位置连续的发生漂移,而且他们的最优方位就大致上以10度/50um的变化绿,按顺时针或逆时针的方向发生连续变化,有时在旋转90~270度以后,旋转方向发生逆转。因此,Hubel和Wiesel设想具有相同最优方位的视皮层细胞,是柱状垂直于皮层表面排列的,而且是连成薄片状的薄层所组成,故如要包括180全方位的这些薄板的总宽度应是0.9mm左右,这样一套薄板结构称为方位柱(orientation column)。眼优势柱:大多数双眼细胞接受双眼输入时,总是有一侧眼占优势的,眼优势决定于交叉和未交叉视通道激活4C层细胞的比例,可以根据分别刺激同侧或对侧眼的感受野所产生反应的大小来决定。眼优势柱与方位柱是相对独立的功能结构系统,他们既不平行又不成直角,而是随即交叉的。空间频率柱,远不如方位柱和眼优势柱那样界限分明。
图13 功能柱
在V2区,构筑物的形状是一些粗细条纹,彼此间被一些轻度染色的中间条纹分开,对波长选择型细胞是集中在细条纹中,对方向性运动有选择性的细胞则是存在于粗条纹中,对形状敏感的细胞则是粗条纹和中间条纹中都有所分布。这些事实是我们可以勾画出四个与不同视觉特征有关的并行系统:一个是针对运动的,一个是针对颜色的,两个是针对形状的。两个计算上大不相同的系统是运动系统和色彩系统,对于运动系统来说,关键性纹状前区是V5,其输入信号的传递是从视网膜经外侧膝状核的大细胞层到V1的4B层,再以直接和通过V2的粗条纹两种方式从那里传送到V5。色彩系统是依靠V4区,其输入信号经外侧膝状核的细胞层传递给V1的色斑,然后以直接和通过V2的细条纹两种方式前往V4。
特定皮层区的损害会相应的造成特有的视觉综合症:V4区的损害曾导致过全色盲(只能看到灰色梯度),不同与单纯色盲,不但不能看见或认识彩色世界,甚至连罹患这种疾病之前所曾见过的色彩是什么样子都回忆不起来。如果它们的视网膜和V1区域都很正常,它们对形状、深度及运动的了解就仍是完好的。V5区的损害会造成运动盲(Akinetopsia),既看不见又不能理解运动中的世界,当物体处于静止状态时,他们可能完全看得见,但是与之相关的运动却会使物体消失,视觉的其他特征却依然没有受损。假定形状和颜色是在皮层内分离的,但是,从来没有人报道过形状是觉得完全丧失和特定丧失。17区不投射到对侧皮层而投射到18,19区和别的皮层区域,18区向前投射到19区,此外,18,19区又投射到对侧皮层对称点和颞叶中部和下部一些区域。
根据以上对视网膜、外膝体和初级视皮层的研究成果,我们可以看到视觉信息处理机制是既平行又分级串行的信息处理机制,如图9。视系统组织成不同的通路对视觉信息的不同侧面进行传递和处理。从外侧膝状体不同层细胞投射至视皮层至少有3条通路,分别对运动、空间(包括立体视觉)信息、形状、图像信息以及颜色信息进行分析处理这三条通路并非完全独立,在不同水平上存在着交叉性连接。单个细胞或细胞群在感知的水平上并不表示某种特征状态,而仅仅表示被感知物的某些特殊的方面。换言之,分离的个部分所表示的并非整体,而使它们间的关系才构成整体的感知。在这种综合的过程中,必须要有“注意”(attention)的参与,“注意”会强调某一物体的特异的性质,突出对生存和生活有重要意义的视觉目标,同时又忽略该物体的其它特性和其他物体。视觉信息的整合是一个过程,在这个过程中对视觉世界的感知和理解是同时发生的。
更高层次的脑信息处理机制,目前提出一些假设,主要有:
1、“主教细胞”假说:根据Hubel和Wiesel提出的分级假说(hierachical hypothesis),就可以推导是否存在祖母细胞。所存在的关键问题是(1)组合爆炸;(2) 信息的集成问题。
2、细胞群假设 :由一群细胞的时空发放模式来反映刺激的类型。问题是(1) 重叠灾难 (2) 难以表达“等级结构”(3)特征捆绑问题。
3、时空编码理论 :动态细胞群。1989年,C.M. Gray & W. Singer等发现了视皮层神经元活动的gamma(40Hz)同步振荡的研究结果,引起了神经科学界的极大关注。1995年,提出了时空编码理论,主要有以下内容:(1) 是建立在神经元脉冲发放的精细时间结构上的,与同步振荡不同的是,脉冲不是等间隔的振荡脉冲,脉冲的间隔也被用于编码信息(2) 神经元是作为脉冲时间一致性检测器来工作的,也就是说,一个神经元自身的脉冲发放情况,由送入该元的输入脉冲在时间上的一致性来确定的,比如说接近同时到达的脉冲数超过一定阈值,则该元发放。与传统神经网络的整合-放电单元不同,通过一致性检测单元传递的不是发放率,而是脉冲发放时间;(3) 通过局部的一致性检测,形成“动态细胞群”。一致性检测是局部事件,动态细胞群则是系统级事件,是一群细胞通过脉冲发放时间临时链接到一起形成的,注意,并不要求这些细胞的脉冲发放是同步的,只要相关就可以了。(4) 相关的关系是根据上下文(任务)形成的,细胞群可以根据上下文的变化进行快速地重组。之所以称为时-空编码,“空”是指空间连接,是一种硬件的拓朴结构,就是神经元间的物理连接;“时”则是指在硬件结构上传递的时间模式,这种模式可以决定动态的连接结构,这种时间模式的产生与硬件连接有关,但时间变量提供了传递信息的巨大能力,结果是可以在同一硬件上形成许多不同的“动态结构”。“动态细胞群”是编码信息的基本单位,是和任务有关的、以自涌方式形成的细胞的瞬时组合。“动态细胞集群”假设可以有效地解决“重叠灾难”问题、捆绑问题、上下文关系表达问题。
对视觉信息处理的根本问题是:
1、 脑的各部分越来越趋向于分工处理各种各样的视觉特征,然后我们的视觉却把各种视觉特征综合成一个完整的视觉实体。脑象——〉心理象、认知象?脑活动——〉精神活动?
2、 视觉图像如何在脑中贮存和利用。在梦中,转动头部探望侧面或背面的物体,视野中的景象在视网膜上的光学象也朝与头部运动相反的方向移动,但看到的外界景象却纹丝不同,脑似乎有补偿机制。