视景医疗
早期视觉环境与视觉功能发育
人类通过视觉系统感知外界缤纷多彩的世界。视觉是脑内最复杂的感觉,人的70%以上的外界信息都是由视觉系统所接收、感知和知觉的。譬如,人的听神经有3万根左右神经纤维,而视神经则多达100万根。从双眼的视网膜(retina)到外膝体(later-algeniculatenucleus,LGN)再到大脑视皮层(V1)的视觉神经通路具有复杂而有序的神经连接模式,而视通路的各级亦存在着精确的视网膜拓扑投射(ret-inotopy)图谱以及处理视觉信息的并行式神经回路。然而人类和哺乳动物视皮层在刚出生时处于尚未发育成熟的状态,从出生后到发育成熟之前的一段时期内,视皮层的神经联系和突触结构能够根据不同的视觉经验(visualexperience)做出适当的调整和改变,早期视觉环境对视觉功能发育有着强有力的影响。
刚出生的婴儿是通过出生后丰富的视觉刺激,逐渐形成复杂的视觉功能。这些视觉通路的畅通是经由亿万个神经细胞有序连接完成的。婴儿和儿童期如果没有丰富的视觉刺激,不但无法正常形成视觉功能,甚至会使大脑视觉潜能完全丧失。出生后6个月开始,视觉功能发育加快,视觉功能成熟在生后的4~5年。所以,从出生开始,孩子们就应该多接触丰富多彩的世界,获得观看各种色彩、物品、景物、现象的视觉机会,以保证良好的视觉功能发育。
1、视觉的中枢通路与皮层定位
视觉通路由4级神经元组成,第一、二、三级神经元位于视网膜内,第四级神经元位于外膝体,由此发出神经纤维最后终止于大脑初级视皮层(V1)。视网膜神经节细胞(retinalganglioncell,RGC,第三级神经元)的轴突组成视神经(opticnerve),离眼球后进入颅腔,在第三脑室底面汇成视交叉(opticchi-asm),约有60%的轴突纤维交叉到对侧,40%的轴突保留在同侧。来自两眼鼻侧视网膜的纤维(即接受颞侧光刺激的部分)都交叉至对侧,并上行至对侧外膝状。而来自颞侧视网膜的纤维(即接受鼻侧光刺激的部分)则不交叉,并上行至同侧外膝体。因此整个视野的左右两半就分别投射至对侧的大脑半球。视神经纤维经视交叉后组成左、右视束(optictract),除了部分到达中脑四叠体(corporaquadri-gemina)参与视调节反射、光反射以及视觉运动反射等活动外,大部分纤维止于外膝体。来自相当于中央凹(foveacentralis)的交叉的视神经纤维终止于外膝体的1、4、6层;不交叉的视神经纤维终止于2、3、5层。因此视觉通路从视网膜开始到视皮层的连接存在精确的视网膜拓扑投射图谱和各种处理视觉信息的并行式神经回路。
2、视觉神经通路的发育
如前所述,视觉通路是从视网膜开始到外膝体,再到视皮层的基本神经连接。出生前视觉系统主要是在基因遗传、分子及内在神经电生理活动的调控下进行发育。出生时视觉系统作为一个整体已基本形成:视觉相关神经元的产生及其功能定位、轴突产生并向下一级目标神经元的投射、视交叉处发生的神经纤维的交叉、视网膜到外膝体和外膝体到初级视皮质的突触连接先后形成、视皮质眼功能柱(ocu-
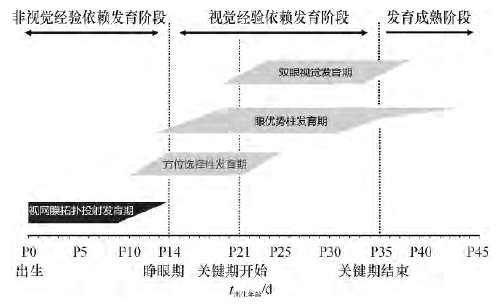
lardominancecolumn)等都已具备和形成。随后这些初始的突触连接将会经历一个精细化的修饰过程(refinement)。在此过程中,神经回路在神经元自发性神经活动(spontaneousneuralactivity)以及由视觉环境和视觉经验调控引导的神经元活动的共同作用下不断进行调整,包括对已有的神经元数量类型功能、突触联系形成和分布、视皮质功能柱进行调节及修饰,逐渐在视觉通路各级产生复杂而有序、有效的拓扑式突触连接体系,最终发育为成熟的视觉神经系统,因此视觉系统的发育是由遗传和环境共同来塑造。从小鼠、猫、猴到人类的不同哺乳动物都经历相似的阶段,即出生后到睁眼期的非视觉经验依赖的拓扑投射的发育阶段以及睁眼后的视觉经验依赖性(visualexperience-dependent)的皮层细胞的感受野(receptivefields)与方位柱(orientationcolumn)的发育、眼优势柱的发育以及双眼视觉(binocularvi-sion)发育等阶段(图1)。
实验证明拓扑投射图谱的形成是由视网膜到视皮层各级细胞信号转导和自发性神经电活动来决定的:视网膜神经节细胞RGC会自发产生有规律的成串动作电位,并募集临近的细胞,进而产生相对同步化的自发性电活动,这种电活动逐渐向周围细胞传播,形成内源性的视网膜波(retinalwaves)。用光学成像的方法观察新生小鼠发现,自发性视网膜活动在睁眼前的整个视觉系统中都存在和传播。改变视网膜活动能够扰乱左右眼的神经节细胞的轴突分离进入各自的眼睛特定图层(eye-specificlayer)。利用光遗传技术(optogenetictechniques)直接操纵视网膜活动发现:神经元同步化活动和短暂的双眼视网膜活动共同调制了视觉系统眼特异性的分离和神经环路的拓扑投射。注射海豚毒素(TTX)阻断视网膜神经节细胞Na+通道或使用地棘蛙素(epi-batidine)作用于nACh受体,能够引起Na+和Ca2+离子通透性增加,去极化而阻止神经节细胞的自发放电,均影响左右眼的神经节细胞轴突自外膝体上分离到各自的眼睛特定图层中以及视皮层眼优势柱的形成。外膝体细胞和视皮层神经元也发现有类似于视网膜波的自发性神经电活动。目前普遍认为,视网膜波对视网膜神经节细胞轴突在外膝体的分层分布也有指导性的作用,在外膝体向视皮层的拓扑投射过程中也扮演有重要角色;与视网膜相似,外膝体细胞的同步化活动则对外膝体到视皮层的拓扑投射的形成具有指导性作用。
(2)视觉经验依赖性的发育阶段:尽管视网膜的神经节细胞到视皮层轴突投射组成了高度精密的点对点(pointtopoint)的连接,但皮层细胞的方位选择性(orientationselectivity)的形成需要睁眼后,视觉信息刺激视网膜最终在皮层上产生视觉反应,没有视觉的刺激和训练,皮层细胞就不能形成方位选择性。成熟视皮层的另一个重要表现是眼优势功能柱,两眼在整个初级视皮层上形成各自占优的区域。视皮层的第4层中相互交织在一起的外膝体投射纤维分离从而形成左右眼各自占优的眼优势柱,由皮层第4层左、右眼优势区域发出的轴突支配到视皮层的上层和下层(第2/3、第5,6层)从而形成双眼视觉的结构基础。双眼视觉是人类最高级的视觉功能,正是因为形成了双眼视觉,人类才能更准确地获得外界物体形状、方位、距离等概念,才能正确判断并适应自身与客观环境间的位置关系。双眼视觉必须是在睁眼后,视觉刺激训练后才能形成,是经验依赖性的发育。由于双眼视觉是一种在动物种属发展晚期获得的本领,同时也是一种非常复杂的生理机制,所以在内、外环境因素的影响下容易遭到破坏而产生紊乱。
早期视觉环境和视觉训练对正常的视觉功能发育起了关键的作用。猫在生后10d前后眼睛睁开,在生后第4~8周猫的视皮层神经细胞功能对视觉刺激和训练最为敏感,实验显示:从猫睁眼开始,每天将小猫置于一个只能看见水平或垂直的筒内5h。5个月后,在垂直条纹环境中长大的小猫只对垂直条纹做出反应,水平条纹环境中长大的小猫只对水平条纹做出反应。然而,眼优势柱的结构形成并不依赖视觉经验,在黑暗中出生并长大的猫或出生后便双眼缝合的猫,其视皮层细胞的眼优势和眼优势柱的发育基本上正常,但大多数不能为双眼驱动,而且其视皮层细胞的方位选择性和方向选择性均明显下降,方位、方向选择性细胞比例亦大为减少。尽管外膝体显示出略弱于皮层的方位选择性,但黑暗中出生并长大的猫,外膝体的方位选择性不受影响,进一步说明皮层的视觉功能发育一定要有视觉的刺激和训练。猫在生后第4~8周,单眼剥夺(monoculardeprivation)导致单眼视觉输入的丧失会造成视皮层的双眼细胞数量大量减少,剥夺眼支配的单眼细胞几乎消失,大多数皮层细胞只对未剥夺眼的刺激有方应,眼优势柱偏向未剥夺眼。此外,本文作者近年的研究表明,哺乳动物视皮层的方位选择性和双眼视觉的发育不仅需要视觉环境和视觉训练,视皮层内不同片层的发育可塑性同样受到视觉环境的影响。接受丘脑LGN传入的第4层的突触可塑性的起止比第2/3层早,呈现一个可塑性发育的“关键期”,而第2/3层则没有关键期,终生具有可塑性,剥夺视觉刺激可使第2/3层的可塑性升高。视觉剥夺(黑暗饲养)阻碍小鼠视皮层第2/3层的中间抑制性神经突触发育成熟,而不影响第4层中的中间抑制性神经突触发育成熟。研究还表明内源性大麻醇及其受体(endocannabinoidsandre-ceptors)在视皮层中间抑制神经元(GABAergicinter-neuron)发育成熟过程中的起重要作用,视皮层的抑制神经元的突触在其靶细胞上的分枝和增殖需要内源性大麻醇及其受体的参与。视觉刺激诱发内源性大麻醇从而导致中间抑制性神经突触发育成熟。这些结果从细胞水平上至少部分解释了在皮层的视觉功能发育过程中视觉刺激和训练的必要性。
以上实验证明:尽管在睁眼前,外膝体发出的来自左右眼睛特定图层的投射纤维在视皮层的第4层的分离形成左右眼各自占优的眼优势柱,但皮层细胞对方位的辨别能力的形成需要早期的视觉刺激。双眼视觉即两眼同时看到同一物体并鉴别形状、方位、距离上的差异的功能需要双眼的同时刺激和训练,在关键期(criticalperiod,CP),如果一只眼丧失视觉刺激,两眼不能在传入信息时间上、信息量上的相互匹配,最终导致完全的单眼视觉,双眼视觉的功能丧失。因此在视觉发育的关键期保证双眼的正常视觉功能非常必要。异常的视觉环境经验会不可逆地导致弱视(amblyopia)等视觉损伤,如视觉刺激剥夺、单眼弱视或斜视(strabismus)、双眼屈光参差(refractiveerror)、高度屈光不正、形觉剥夺等异常的视觉经验都会造成视觉功能异常。猴在刚出生时视觉系统就已相当成熟,其关键期是从出生开始一直延续到生后第6个月,在第6~8周最敏感。小鼠和大鼠的视皮层发育关键期在睁眼后开始持续到生后第5周,以生后28~32d最为敏感。人类的视觉系统发育的关键期是在1~6岁期间。总之,不同动物的视觉发育关键期时间不同,在视觉发育的关键期内,异常的视觉环境经验会不可逆地导致弱视等视觉损伤。
3、视觉训练的必要性对人类的启示以及重新激活皮层可塑性用于改善弱视等视觉功能异常的尝试
明确了视觉神经通路的非视觉经验和视觉经验依赖的发育过程,特别是视觉经验对视皮层神经元形成方位选择性和双眼视觉等正常功能的必要性,就可以采取措施来保障我们的视觉发育。婴儿出生后,视觉系统的神经结构作为一个整体已经基本形成,但需要生后的一段时期的视觉训练才能使视觉功能达到成熟。这个时期就是人类视觉发育的关键期,即生后1~6岁。在这个时期内给婴儿足够和丰富的视觉刺激,孩子应该获得观看各种色彩、物品、景物、现象的机会来保证正常的视觉功能。此外儿童如有视觉异常,如两眼屈光不齐、斜视或弱视等,都要尽可能在视觉发育关键期内进行治疗而纠正。处于关键期的治疗,才有可能使视功能得到提高,而在成人后却很难得到改善,主要是因为成人已过了视觉发育的关键期,神经环路很难再被视觉经验所塑造。
近10年来,研究人员试图重新激活视皮层发育的可塑性,即运用神经药理学、细胞移植、基因调控等方法等侵入式的途径来激活关键期,以探寻改善弱视等视觉功能障碍的有效治疗手段。研究表明发现,导致视皮层发育关键期关闭的因素是:(1)视皮层中抑制性神经元逐渐发育成熟,并导致环路中的兴奋和抑制达到平衡(I/E)而终止关键期;(2)神经元细胞外基质发育成熟,从而限制并固化了神经回路,最终使关键期结束。因此研究者通过(1)施用影响GABA合成和释放或阻断GABA受体的药物,降低皮层内神经元抑制性强度,打破I/E平衡;(2)利用细胞外基质相关的水解酶、抑制性GABA神经元的移植以及特定基因敲除的方法,降低发育成熟的细胞外基质对神经元的固化作用,使皮层内神经网路松绑这两种主要方法来重新激活皮层发育可塑性,但通过药物或改变基因等侵入性的方式,其在临床上医用上的高风险和多限制仍然是一个难以解决的问题。
近年来,非侵入式(non-invasive)的方法来重新激活视皮层可塑性的尝试受到更多青睐。如让弱视病人参视觉认知学习强化训练,要求仅用弱视眼完成一系列复杂的视觉相关任务,经过一定的训练疗程后,病人的弱视眼功能较未训练病人得到持久改善;采用动作视频游戏的方法,可提高弱视病人包括视觉认知、视觉-动作协调能力、空间认知、注意力等综合视觉能力,有效地重塑成年大脑皮层神经回路进而恢复正常视觉功能。此外,Sale等将成年动物经过丰富环境饲养(environmentalenrichment,EE),也可以使之在关键期之后恢复视皮层的可塑性。非侵入的方法提高可塑性的具体机制仍然不是很清楚,可能涉及到运动、感觉、听觉、视觉等多个皮层之间跨模态可塑性,以及社会活动所触发的情绪、抉择、注意力等复杂认知神经活动,大脑通过有效分散和重置机体的神经递质系统和神经调质系统,综合降低皮层内的抑制强度(重置I/E平衡)或软化皮质中的神经元周围基质网络(PNNs),从而重新激活皮层的可塑性。
4、评价与展望
大脑发育的早期阶段,神经环路会经历复杂而有序的发生发育过程。50多年来对视觉系统拓扑投射图谱与视皮层神经环路的基本构建程式进行了深入的研究,对特定自发性神经电活动与视觉经验影响视觉通路的机制有了初步理解。尽管早期的视觉环境和经验对成熟的视觉功能非常必要,但早期的视觉环境和视觉训练是如何引起视皮层内部神经回路成为有功能的图式的机制仍然不清。
在视觉发育的关键期内若受到斜视、屈光不正等异常的视觉环境经验,儿童的视觉系统发育就会受到影响,由于视觉发育的关键期只存在于幼年时期,错过了这个时期弱视就比较难矫正。近期的研究表明,饮食、睡眠也分别对增强和巩固视皮层的可塑性有重要作用,因此应综合考虑并改善儿童早期生活环境,才能使他们的视觉系统得到最完善的发育,使他们的大脑可塑性达到最高的水平。近来研究者尝试用非侵入式(non-invasive)的方法来重新激活皮层可塑性,如治疗成人弱视病人等。丰富环境对激活动物皮层可塑性非常有效,它可以使得神经回路发生分子,解剖和功能上的改变;加速视皮层功能的发育、特别是可以完全抵消暗室饲养导致视皮层功能的改变,但如何在人群中实施丰富环境的方法来改善或矫正视觉功能,有待将来的临床实践。对成年的弱视病人用弱视眼完成一系列复杂的视觉相关任务以及让弱视眼完成动作视频游戏的方法值得推荐,在临床上也有可行性。弱视病人采用上述方法,经过一定的训练之后,弱视眼视觉功能有望得到持久的改善。
刚出生的婴儿是通过出生后丰富的视觉刺激,逐渐形成复杂的视觉功能。这些视觉通路的畅通是经由亿万个神经细胞有序连接完成的。婴儿和儿童期如果没有丰富的视觉刺激,不但无法正常形成视觉功能,甚至会使大脑视觉潜能完全丧失。出生后6个月开始,视觉功能发育加快,视觉功能成熟在生后的4~5年。所以,从出生开始,孩子们就应该多接触丰富多彩的世界,获得观看各种色彩、物品、景物、现象的视觉机会,以保证良好的视觉功能发育。
1、视觉的中枢通路与皮层定位
视觉通路由4级神经元组成,第一、二、三级神经元位于视网膜内,第四级神经元位于外膝体,由此发出神经纤维最后终止于大脑初级视皮层(V1)。视网膜神经节细胞(retinalganglioncell,RGC,第三级神经元)的轴突组成视神经(opticnerve),离眼球后进入颅腔,在第三脑室底面汇成视交叉(opticchi-asm),约有60%的轴突纤维交叉到对侧,40%的轴突保留在同侧。来自两眼鼻侧视网膜的纤维(即接受颞侧光刺激的部分)都交叉至对侧,并上行至对侧外膝状。而来自颞侧视网膜的纤维(即接受鼻侧光刺激的部分)则不交叉,并上行至同侧外膝体。因此整个视野的左右两半就分别投射至对侧的大脑半球。视神经纤维经视交叉后组成左、右视束(optictract),除了部分到达中脑四叠体(corporaquadri-gemina)参与视调节反射、光反射以及视觉运动反射等活动外,大部分纤维止于外膝体。来自相当于中央凹(foveacentralis)的交叉的视神经纤维终止于外膝体的1、4、6层;不交叉的视神经纤维终止于2、3、5层。因此视觉通路从视网膜开始到视皮层的连接存在精确的视网膜拓扑投射图谱和各种处理视觉信息的并行式神经回路。
2、视觉神经通路的发育
如前所述,视觉通路是从视网膜开始到外膝体,再到视皮层的基本神经连接。出生前视觉系统主要是在基因遗传、分子及内在神经电生理活动的调控下进行发育。出生时视觉系统作为一个整体已基本形成:视觉相关神经元的产生及其功能定位、轴突产生并向下一级目标神经元的投射、视交叉处发生的神经纤维的交叉、视网膜到外膝体和外膝体到初级视皮质的突触连接先后形成、视皮质眼功能柱(ocu-
lardominancecolumn)等都已具备和形成。随后这些初始的突触连接将会经历一个精细化的修饰过程(refinement)。在此过程中,神经回路在神经元自发性神经活动(spontaneousneuralactivity)以及由视觉环境和视觉经验调控引导的神经元活动的共同作用下不断进行调整,包括对已有的神经元数量类型功能、突触联系形成和分布、视皮质功能柱进行调节及修饰,逐渐在视觉通路各级产生复杂而有序、有效的拓扑式突触连接体系,最终发育为成熟的视觉神经系统,因此视觉系统的发育是由遗传和环境共同来塑造。从小鼠、猫、猴到人类的不同哺乳动物都经历相似的阶段,即出生后到睁眼期的非视觉经验依赖的拓扑投射的发育阶段以及睁眼后的视觉经验依赖性(visualexperience-dependent)的皮层细胞的感受野(receptivefields)与方位柱(orientationcolumn)的发育、眼优势柱的发育以及双眼视觉(binocularvi-sion)发育等阶段(图1)。
小鼠睁眼之前(生后 14 d,P14),从视网膜到初级视皮层的基本拓扑投射已经形成。睁眼后,方位选择性以及对侧眼所驱动的神经元活动加强;皮层关键期开始的早期(P20 -25),皮层神经元的感受野与方位柱等结构形成,随后,神经元的视觉反应和对同侧眼的响应显著增加,眼优势柱逐渐形成,最终形成双眼视觉。关键期结束后(P33 -35),视觉通路完全发育成熟、视觉功能完善。视网膜到视皮层的拓扑投射的形成不需要视觉经验,然而,视皮层细胞感受野与方位柱、眼优势柱及双眼视觉等功能发育是视觉经验依赖性的发育.
图1:哺乳动物(小鼠)视觉系统的发育过程
Fig.1:Development of mammalian (mouse) visual system
(1)非视觉经验依赖的拓扑投射的发育阶段:大鼠,小鼠以及猫在视网膜神经节细胞被视杆(rodcells)和视锥(conecells)光感受细胞(photoreceptorcells)驱动之前,视网膜的神经节细胞的轴突投射到背侧外膝体。一开始,来自不同眼的神经节细胞的轴突在外膝体上交织重叠分布,然后相互分离进入各自的眼睛特定图层(eye-specificlayer)中。猫的外膝体为3层构造,来自对侧眼的视网膜神经节细胞轴突终止于A层和C层、同侧眼的纤维却止于A1层。在胚胎发育的末期(E18-21),大鼠和小鼠的来自左右眼视网膜神经节纤维均已投射到外膝体,但在外膝体上交错、重叠分布。在后来的轴突精细化过程中,约在生后的第1周(P1~8)来自同侧眼的纤维大部分被删减(elimination),在外膝体上占有的领域很小,而对侧纤维则得到加强和增多,占有较大的领域,形成了啮齿类动物对侧眼占绝对优势的视觉功能。随后,从外膝体发出的来自眼睛特定图层的神经轴突纤维投射到视皮层的第4层(layer4)上,相互重叠地和视皮层第4层的神经建立突触连接,随后同样发生轴突精细化,原先在视皮层第4层中相互重叠的外膝体投射纤维相互分离,从而形成不同眼睛特定图层的支配区,即左右眼各自支配的眼优势柱。猴和人在出生时外膝体的眼睛特定图层和视皮层的眼优势柱的神经结构已经形成,说明灵长类基本的视觉神经通路在胚胎期已发育相对完好。到此为止,从视网膜的神经节细胞到视皮层轴突投射组成了高度精密的点对点的神经连接,视觉通路逐渐形成了各级复杂而有序的拓扑式突触连接体系,视觉通路的环路结构得以完善。这个发育阶段是动物在睁眼前完成的,不需要视觉经验。图1:哺乳动物(小鼠)视觉系统的发育过程
Fig.1:Development of mammalian (mouse) visual system
实验证明拓扑投射图谱的形成是由视网膜到视皮层各级细胞信号转导和自发性神经电活动来决定的:视网膜神经节细胞RGC会自发产生有规律的成串动作电位,并募集临近的细胞,进而产生相对同步化的自发性电活动,这种电活动逐渐向周围细胞传播,形成内源性的视网膜波(retinalwaves)。用光学成像的方法观察新生小鼠发现,自发性视网膜活动在睁眼前的整个视觉系统中都存在和传播。改变视网膜活动能够扰乱左右眼的神经节细胞的轴突分离进入各自的眼睛特定图层(eye-specificlayer)。利用光遗传技术(optogenetictechniques)直接操纵视网膜活动发现:神经元同步化活动和短暂的双眼视网膜活动共同调制了视觉系统眼特异性的分离和神经环路的拓扑投射。注射海豚毒素(TTX)阻断视网膜神经节细胞Na+通道或使用地棘蛙素(epi-batidine)作用于nACh受体,能够引起Na+和Ca2+离子通透性增加,去极化而阻止神经节细胞的自发放电,均影响左右眼的神经节细胞轴突自外膝体上分离到各自的眼睛特定图层中以及视皮层眼优势柱的形成。外膝体细胞和视皮层神经元也发现有类似于视网膜波的自发性神经电活动。目前普遍认为,视网膜波对视网膜神经节细胞轴突在外膝体的分层分布也有指导性的作用,在外膝体向视皮层的拓扑投射过程中也扮演有重要角色;与视网膜相似,外膝体细胞的同步化活动则对外膝体到视皮层的拓扑投射的形成具有指导性作用。
(2)视觉经验依赖性的发育阶段:尽管视网膜的神经节细胞到视皮层轴突投射组成了高度精密的点对点(pointtopoint)的连接,但皮层细胞的方位选择性(orientationselectivity)的形成需要睁眼后,视觉信息刺激视网膜最终在皮层上产生视觉反应,没有视觉的刺激和训练,皮层细胞就不能形成方位选择性。成熟视皮层的另一个重要表现是眼优势功能柱,两眼在整个初级视皮层上形成各自占优的区域。视皮层的第4层中相互交织在一起的外膝体投射纤维分离从而形成左右眼各自占优的眼优势柱,由皮层第4层左、右眼优势区域发出的轴突支配到视皮层的上层和下层(第2/3、第5,6层)从而形成双眼视觉的结构基础。双眼视觉是人类最高级的视觉功能,正是因为形成了双眼视觉,人类才能更准确地获得外界物体形状、方位、距离等概念,才能正确判断并适应自身与客观环境间的位置关系。双眼视觉必须是在睁眼后,视觉刺激训练后才能形成,是经验依赖性的发育。由于双眼视觉是一种在动物种属发展晚期获得的本领,同时也是一种非常复杂的生理机制,所以在内、外环境因素的影响下容易遭到破坏而产生紊乱。
早期视觉环境和视觉训练对正常的视觉功能发育起了关键的作用。猫在生后10d前后眼睛睁开,在生后第4~8周猫的视皮层神经细胞功能对视觉刺激和训练最为敏感,实验显示:从猫睁眼开始,每天将小猫置于一个只能看见水平或垂直的筒内5h。5个月后,在垂直条纹环境中长大的小猫只对垂直条纹做出反应,水平条纹环境中长大的小猫只对水平条纹做出反应。然而,眼优势柱的结构形成并不依赖视觉经验,在黑暗中出生并长大的猫或出生后便双眼缝合的猫,其视皮层细胞的眼优势和眼优势柱的发育基本上正常,但大多数不能为双眼驱动,而且其视皮层细胞的方位选择性和方向选择性均明显下降,方位、方向选择性细胞比例亦大为减少。尽管外膝体显示出略弱于皮层的方位选择性,但黑暗中出生并长大的猫,外膝体的方位选择性不受影响,进一步说明皮层的视觉功能发育一定要有视觉的刺激和训练。猫在生后第4~8周,单眼剥夺(monoculardeprivation)导致单眼视觉输入的丧失会造成视皮层的双眼细胞数量大量减少,剥夺眼支配的单眼细胞几乎消失,大多数皮层细胞只对未剥夺眼的刺激有方应,眼优势柱偏向未剥夺眼。此外,本文作者近年的研究表明,哺乳动物视皮层的方位选择性和双眼视觉的发育不仅需要视觉环境和视觉训练,视皮层内不同片层的发育可塑性同样受到视觉环境的影响。接受丘脑LGN传入的第4层的突触可塑性的起止比第2/3层早,呈现一个可塑性发育的“关键期”,而第2/3层则没有关键期,终生具有可塑性,剥夺视觉刺激可使第2/3层的可塑性升高。视觉剥夺(黑暗饲养)阻碍小鼠视皮层第2/3层的中间抑制性神经突触发育成熟,而不影响第4层中的中间抑制性神经突触发育成熟。研究还表明内源性大麻醇及其受体(endocannabinoidsandre-ceptors)在视皮层中间抑制神经元(GABAergicinter-neuron)发育成熟过程中的起重要作用,视皮层的抑制神经元的突触在其靶细胞上的分枝和增殖需要内源性大麻醇及其受体的参与。视觉刺激诱发内源性大麻醇从而导致中间抑制性神经突触发育成熟。这些结果从细胞水平上至少部分解释了在皮层的视觉功能发育过程中视觉刺激和训练的必要性。
以上实验证明:尽管在睁眼前,外膝体发出的来自左右眼睛特定图层的投射纤维在视皮层的第4层的分离形成左右眼各自占优的眼优势柱,但皮层细胞对方位的辨别能力的形成需要早期的视觉刺激。双眼视觉即两眼同时看到同一物体并鉴别形状、方位、距离上的差异的功能需要双眼的同时刺激和训练,在关键期(criticalperiod,CP),如果一只眼丧失视觉刺激,两眼不能在传入信息时间上、信息量上的相互匹配,最终导致完全的单眼视觉,双眼视觉的功能丧失。因此在视觉发育的关键期保证双眼的正常视觉功能非常必要。异常的视觉环境经验会不可逆地导致弱视(amblyopia)等视觉损伤,如视觉刺激剥夺、单眼弱视或斜视(strabismus)、双眼屈光参差(refractiveerror)、高度屈光不正、形觉剥夺等异常的视觉经验都会造成视觉功能异常。猴在刚出生时视觉系统就已相当成熟,其关键期是从出生开始一直延续到生后第6个月,在第6~8周最敏感。小鼠和大鼠的视皮层发育关键期在睁眼后开始持续到生后第5周,以生后28~32d最为敏感。人类的视觉系统发育的关键期是在1~6岁期间。总之,不同动物的视觉发育关键期时间不同,在视觉发育的关键期内,异常的视觉环境经验会不可逆地导致弱视等视觉损伤。
3、视觉训练的必要性对人类的启示以及重新激活皮层可塑性用于改善弱视等视觉功能异常的尝试
明确了视觉神经通路的非视觉经验和视觉经验依赖的发育过程,特别是视觉经验对视皮层神经元形成方位选择性和双眼视觉等正常功能的必要性,就可以采取措施来保障我们的视觉发育。婴儿出生后,视觉系统的神经结构作为一个整体已经基本形成,但需要生后的一段时期的视觉训练才能使视觉功能达到成熟。这个时期就是人类视觉发育的关键期,即生后1~6岁。在这个时期内给婴儿足够和丰富的视觉刺激,孩子应该获得观看各种色彩、物品、景物、现象的机会来保证正常的视觉功能。此外儿童如有视觉异常,如两眼屈光不齐、斜视或弱视等,都要尽可能在视觉发育关键期内进行治疗而纠正。处于关键期的治疗,才有可能使视功能得到提高,而在成人后却很难得到改善,主要是因为成人已过了视觉发育的关键期,神经环路很难再被视觉经验所塑造。
近10年来,研究人员试图重新激活视皮层发育的可塑性,即运用神经药理学、细胞移植、基因调控等方法等侵入式的途径来激活关键期,以探寻改善弱视等视觉功能障碍的有效治疗手段。研究表明发现,导致视皮层发育关键期关闭的因素是:(1)视皮层中抑制性神经元逐渐发育成熟,并导致环路中的兴奋和抑制达到平衡(I/E)而终止关键期;(2)神经元细胞外基质发育成熟,从而限制并固化了神经回路,最终使关键期结束。因此研究者通过(1)施用影响GABA合成和释放或阻断GABA受体的药物,降低皮层内神经元抑制性强度,打破I/E平衡;(2)利用细胞外基质相关的水解酶、抑制性GABA神经元的移植以及特定基因敲除的方法,降低发育成熟的细胞外基质对神经元的固化作用,使皮层内神经网路松绑这两种主要方法来重新激活皮层发育可塑性,但通过药物或改变基因等侵入性的方式,其在临床上医用上的高风险和多限制仍然是一个难以解决的问题。
近年来,非侵入式(non-invasive)的方法来重新激活视皮层可塑性的尝试受到更多青睐。如让弱视病人参视觉认知学习强化训练,要求仅用弱视眼完成一系列复杂的视觉相关任务,经过一定的训练疗程后,病人的弱视眼功能较未训练病人得到持久改善;采用动作视频游戏的方法,可提高弱视病人包括视觉认知、视觉-动作协调能力、空间认知、注意力等综合视觉能力,有效地重塑成年大脑皮层神经回路进而恢复正常视觉功能。此外,Sale等将成年动物经过丰富环境饲养(environmentalenrichment,EE),也可以使之在关键期之后恢复视皮层的可塑性。非侵入的方法提高可塑性的具体机制仍然不是很清楚,可能涉及到运动、感觉、听觉、视觉等多个皮层之间跨模态可塑性,以及社会活动所触发的情绪、抉择、注意力等复杂认知神经活动,大脑通过有效分散和重置机体的神经递质系统和神经调质系统,综合降低皮层内的抑制强度(重置I/E平衡)或软化皮质中的神经元周围基质网络(PNNs),从而重新激活皮层的可塑性。
4、评价与展望
大脑发育的早期阶段,神经环路会经历复杂而有序的发生发育过程。50多年来对视觉系统拓扑投射图谱与视皮层神经环路的基本构建程式进行了深入的研究,对特定自发性神经电活动与视觉经验影响视觉通路的机制有了初步理解。尽管早期的视觉环境和经验对成熟的视觉功能非常必要,但早期的视觉环境和视觉训练是如何引起视皮层内部神经回路成为有功能的图式的机制仍然不清。
在视觉发育的关键期内若受到斜视、屈光不正等异常的视觉环境经验,儿童的视觉系统发育就会受到影响,由于视觉发育的关键期只存在于幼年时期,错过了这个时期弱视就比较难矫正。近期的研究表明,饮食、睡眠也分别对增强和巩固视皮层的可塑性有重要作用,因此应综合考虑并改善儿童早期生活环境,才能使他们的视觉系统得到最完善的发育,使他们的大脑可塑性达到最高的水平。近来研究者尝试用非侵入式(non-invasive)的方法来重新激活皮层可塑性,如治疗成人弱视病人等。丰富环境对激活动物皮层可塑性非常有效,它可以使得神经回路发生分子,解剖和功能上的改变;加速视皮层功能的发育、特别是可以完全抵消暗室饲养导致视皮层功能的改变,但如何在人群中实施丰富环境的方法来改善或矫正视觉功能,有待将来的临床实践。对成年的弱视病人用弱视眼完成一系列复杂的视觉相关任务以及让弱视眼完成动作视频游戏的方法值得推荐,在临床上也有可行性。弱视病人采用上述方法,经过一定的训练之后,弱视眼视觉功能有望得到持久的改善。